Azawakhs de Garde-Épée
- Accueil
- L'Azawakh
- Nos Azawakhs
- Les Chiots
- Les portées
 Blog
Blog- Contact

![]()
En été 2002, quelques membres du comité directeur du SLAG français avaient proposé d'améliorer la base reproductrice des Azawakhs par une retrempe ciblée avec des Sloughis. Le potentiel génétique de l'Azawakh en Europe serait trop restreint. Cette crainte est-elle justifiée? Pour quelles raisons? Qu'est-il possible d'entreprendre? Avant de répondre à ces questions, j'évoquerais les bases pertinentes de la génétique biologique sous la forme la plus simple. La base: La cellule La cellule se compose de l'enveloppe et du noyau. Celui-ci est le porteur du matériel génétique. Il contient des filaments centraux ou les chromosomes qui existent toujours par paires. Le chien possède donc 78 chromosomes, 39 couples. Le matériel de transmission (DNS) est organisé sous forme de sections. Ils sont désignés comme des microsatellites et sont des marqueurs génétiques. Chez le chien, il y a jusqu'à 50.000 microsatellites. Chaque microsatellite se tient à une place bien précise sur un chromosome et est différent d'un chien à l'autre. Plus deux chiens sont étroitement apparentés, plus leurs microsatellites sont semblables. Les gènes en tant que responsables de la transmission héréditaire Les plus petites unités de l'information génétique sont les gènes. Puisque les chromosomes n'esistent pas individuellemnt mais par paires - de manière homologue -, chaque gène posséde un gène partenaire ou homologue (allèle).
On considère que les chiens ont plus de 100.000 couples d'allèle, donc environ 200.000 gènes. La section, sur laquelle deux allèles sont localisés sur un chromosome, s'appelle une place génétique (locus). Un gène spécifique apparaît toujours à la même place du chromosome. Et ce gène spécifique influencera toujours une qualité tout à fait précise et seulement celle-ci. Puisqu'un chromosome du couple de chromosomes provient du père et l'autre de la mère, la moitié de tous les gènes proviennent de l'étalon et l'autre moitié de la chienne. Un
allèle peut dominer l'autre et ainsi
déterminer ce qui sera transmis. Si
un tel allèle partage un locus avec
un allèle plus faible, l'allèle
plus faible est opprimé et l'aspect
du chien (le phénotype) est déterminé
par le gène plus fort et dominant.
Les gènes, qui dans ce cas ne peuvent
s'exprimer, sont désignés par
le terme récessif. Des caractéristiques
récessives deviennent seulement visibles
sur un animal lorsque leurs responsables génétiques
existent de façon "pure",
c.-à-d. par paires sur la même
place génétique (locus) de l'animal.
Cela joue un rôle important dans la
transmission des tares génétiques,
puisque de nombreuses tares sont transmises
de façon récessive. Règles de la transmission génétique L'action
des facteurs génétiques induit
les possibilités suivantes: Pour de nombreuses caractèristiques un seul gène spécifique n'est pas responsable; plusieurs gènes différents agissent ensembles. De tels transmissions polygéniques existent pour presque toutes les qualités et caractéristiques quantitatives. Ces dernières sont par exemple la taille, le poids, l'angulation arrière, les qualités de performance ou de tempéremment. Les caractéristiques qualitatives sont le poil ou la robe. Certains gènes actifs ne se font remarquer que lorsqu'ils apparaissent dans une minorité d'animaux. De tels effets apparaissent aussi au niveau des tares génétiques. Certaines tares sont des modifications (mutations) de gènes sains qui se transmettent généralement de façon récessive. Elles se transmettent souvent sur des générations dans l'obscurité. Ce n'est que lors de la réunion de deux de ces gènes mutants que l'on peut voir surgir de manière perceptible ceux-ci dans le phénotype. L'Héritabilité
est l'influençabilité du phénotype
par le génotype. Elle peut varier de
0 à 100 pour cent. Une qualité
est rarement transmissible à 100 pour
cent (cf. Claude Gaillard, Notre chien de
race, décembre 2002).
illustration
2 (: Hansen, Hérédité
chez le chien, 2001)
La
consanguinité est un moyen visant à
obtenir des caractéristiques ou qualités
souhaitées ou au contraire à
éliminer des caractéristiques
et qualités indésirables. La
règle de base simple, selon toute apparence,
de l'élevage est que seuls les sujets
qui correspondent les plus dans leurs caractéristiques
et qualités à l'objectif d'élevage
sont utilisés comme reproducteurs -
la question qui reste naturellement ouverte,
est s'il existe un progrès d'élevage,
c.-à-d. si un programme d'élevage
choisi a réellemnt un sens. D'habitude, on peut recourir en élevage à un large choix de sujets reproducteurs non apparentés, lorsque des défauts apparaissent avec une sélection homozygote et doivent être corrigés. Cette possibilité est fort réduite lorsqu'on a à faire avec une population numériquement limitée comme chez l'Azawakhs. Car la sélection signifie toujours une modification de fréquence génétique et/ou des pertes génétiques, c.-à-d. qu'avec une sélection ciblée et toujours réitérée la fréquence de l'apparition de certains gènes dans la population reproductrice totale change. Pour l'estimation des pertes génétiques, l'évaluation d'un reproducteur, au moyen de son coefficient de consanguinité et de son coefficient de perte g Le
coefficient de consanguinité d'un chien
indique, combien l'Hétérozygotie
a perdu et l'Homozygotie a gagné en
pourcentage par rapport à la moyenne
de la race. Les
coefficients de perte génétique
(AVK) (formule du Prof. Schlegel, université
Vienne) est le quotient du nombre d'ancêtres
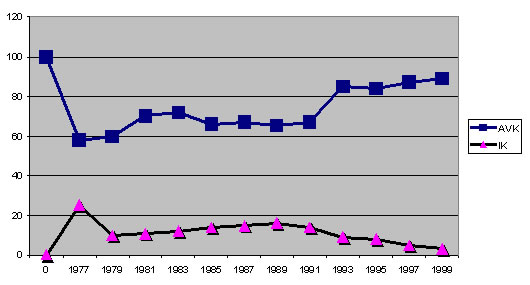
uniques et du nombre total des ancêtres. En ce qui concerne la question d'entrée "appauvrissement génétique de l'Azawakh?" j'ai analysé la population européenne (environ 900 sujets de la période de 1977 à 2001) concernant l'IK et l'AVK. Pour obtenir des valeurs moyennes annuelles, toutes les valeurs IK et AVK des sujets documentées ont été calculées , ces valeurs ont été ajoutées respectivement et divisées par le nombre des entrées. Le résultat pour la population d'Azawakhs en France montre la diminution constante de l'AVK jusqu'en dessous de la valeur de tolérance génétique de 70 pour cent à la fin des années quatre-vingt-dix. Le coefficient de consanguinité augmente proportionnellement au-dessus du seuil critique de 20 pour cent. Cela donne un "effet de col de bouteille" de l'éventail génétique initial des Azawakhs importés d'Afrique coloniale à un rétrécissement dramatique qui n'est plus corrigible avec le potentiel génétique du cheptel reproducteur français actuel. L'élevage Allemand a atteint dans les années soixante-dix un stade qui a plus ou moins correspondu à l'état actuel de la population française. Par l'apport d'Azawakhs en provenance de France et surtout depuis le recours à des importations d'Afrique depuis les années quatre-vingt-dix, l'élevage en général en Allemagne retrouve des valeurs croissantes et sures. Certains élevages et lignées particulières peuvent dévier naturellement plus ou moins fortement de cette moyenne statistique (cf. illustrations 6 et 8). Répartition IK et AVK de l'élevage français: Répartition
IK et AVK de l'élevage allemand: L'Homozygotie est utilisée en élevage d'animaux de compagnie comme moyen d'amélioration des races. La consanguinité vise à produire des chiens au potentiel génétique homozygote. Plus IK est élevé, plus grande est la probabilité que les descendants soient homozygotes pour certains gènes. Il reste que l'on peut se demander quels gènes deviennent homozygotes lors de ces acouplements, seuls les gènes souhaités ou aussi les indésirables. Chez une population d'élevage déjà faible et restreinte génétiquement cela peut conduire à l'effondrement d'une race. Les tares héréditaires sont habituellement déterminées par des gènes récessifs transmis généralement de façon polygénique. La plupart des qualités ou caractéristiques indésirables qui apparaissent au cours de sélections homozygotes en ligne (linebreeding), sont des tares héréditaires. Seuls les descendants de reproducteurs qui possèdent le gène récessif de la tare de façon homozygote afficheront ce phénotype. Ainsi, il est possible qu'une certaine caractéristique souhaitée qui devait être fixée par consanguinité, soit couplée avec une autre caractéristique indésirable. Ou bien les gènes que l'on veut voir disparaître par consanguinité sont couplés avec une autre caractéristique particulièrement recherchée. Au sein de petits élevages des conséquences positives et négatives apparaissent déjà à court terme avec la consanguinité. Lors de la reconstruction d'une population reproductrice, la consanguinité peut renforcer relativement rapidement des caractéristiques raciales définies par les éleveurs ou les fédérations (souvent tout à fait arbitrairement). En augmentant la consanguinité apparaissent alors certains problèmes que l'on qualifie de dépression génétique. Quelques
conséquences de la consanguinité
(caractéristiques de dépression
génétique)
illustration
5 ( Hansen, Hérédité
chez le chien ; 2001)
Les conséquences négatives de la consanguinité n'apparaissent pas immédiatement ou toutes ensemble. Certaines lignées semblent tolérer un degré de consanguinité plus élevé que d'autres, avant que des effets négatifs ne se fassent sentir. Même au sein d' une sélection en ligne étroite avec des IK et AVK relativement élevés, des tares génétiques peuvent rester cachées sur de longues périodes. Elles peuvent, par de heureux hasards génétiques, rester confinées et/ou ne concerner que certaines qualités physiques ou psychiques qui sont moins visibles ou sont acceptées tacitement par les éleveurs et les propriétaires. De cette façon même, naît le danger que des conséquences négatives sur l'aspect et le comportement soient, avec le temps, interprétées à tort par les juges et le public comme des caractéristiques naturelles et/ou générale de la race . Par conséquent, l'éventail de la "philosophie d'éleveur" est étonnamment grande. Les IK- et les AVK- des portées de trois élevages d'Azawakhs peuvent servir d'exemple. Répartition
des IK- et AVK- d'un élevage pratiquant
consanguinité et inceste: Répartition
des IK- et AVK- d'un élevage ne pratiquant
pas de consanguinité: Répartition
des IK- et AVK- d'un élevage pratiquant
la sélection en ligne (linbreeding): La consanguinité ne doit pas inéluctablement conduire à la catastrophe, d'autant moins si des acouplements hors consanguinité réalisés à temps, améliorent les IK et AVK. Les seuils de tolérance admis sont un IK: moins de 10 et un AVK: plus de 75. La consanguinité (large ou étroite) signifie toutefois, dans tout les cas, des pertes génétiques irrémédiables et le risque de modifications indésirables et/ou de maladies génétiques. Je suppose que le club français était conscient de ces dangers (illustration 5). Alors, si le diagnostic est correct, la proposition de thérapie, la retrempe avec des Sloughis en guise d'élargissement génétique, aurait toutefois été erronée. Comme moyen de choix, s'offre, comme pratiqué entretemps avec succès dans l'élevage allemand (cf. par exemple illustration 7), le recours au potentiel génétique originel des Azawakhs disponible comme auparavant. |